We often refer to the roles of food in terms of its nutritional function—providing nutrients required for the construction of organs and body components—and its protective role, such as supporting detoxification and anti-aging processes. These functions are mainly attributed to low-molecular-weight compounds that are absorbed into the body. However, food has another important function, namely the ability to stimulate or enhance the immune system.
The immunostimulatory effects of food arise from its ability to counteract exogenous factors invading the body. In food, high-molecular-weight components such as polysaccharides (starch, glucan, and pectin) or proteins, when absorbed, are recognized as foreign or potentially harmful entities or waste products that must be processed and cleared. This recognition serves as a trigger for activation and enhancement of immune responses. Several polyglycosides, including β-glucan, alkyl polyglycosides, gellan gum, and chitosan, have been reported to possess immunostimulatory activities [1, 2].
The immune mechanism can be described as follows: circulating leukocytes patrol the bloodstream to detect abnormal cells/factors. They may capture, engulf, and degrade bacteria or toxic chemicals, while releasing inflammatory mediators such as histamine and cytokines to stimulate the production of immune cells and recruit them to sites of invasion to eliminate foreign factors. Mast cells, upon recognizing exogenous factors (bacteria or toxic chemicals), also secrete histamine to further stimulate the immune system [3].
Bounous Gustavo reported that different protein diets—including lactalbumin (L), casein (C), soybean protein (S), wheat protein (W), and Purina chow (a standard animal feed)—exert distinct effects on humoral immune responses in mice. The immune response of mice fed with diet L was the highest, nearly five-fold higher than those fed diet C. In contrast, responses in groups fed diets C, S, and W were significantly lower than in mice fed the standard chow. The author concluded that the main factor underlying the observed immune effects was not the availability or concentration of individual essential amino acids, but rather the composite effect of the specific amino acid distribution within the protein [4]. In other words, proteins as a whole, rather than single amino acids, appear to exert immunomodulatory effects. This is consistent with the known immune mechanisms against exogenous invasion. It has long been recognized that small amounts of soluble proteins absorbed via the gastrointestinal tract and transported to the periphery are sufficient to elicit immune responses [5].
In food, easily absorbed nutrients are typically low-molecular-weight compounds such as sugars and amino acids, etc. which are absorbed via active transport across the intestinal epithelium. High-molecular-weight compounds or polymer chains can not freely diffuse across the intestinal membrane; however, a small fraction of these can still enter the body via paracellular transport, endocytotic vesicles, or phagocytosis mediated by specialized intestinal M cells [6].
    |
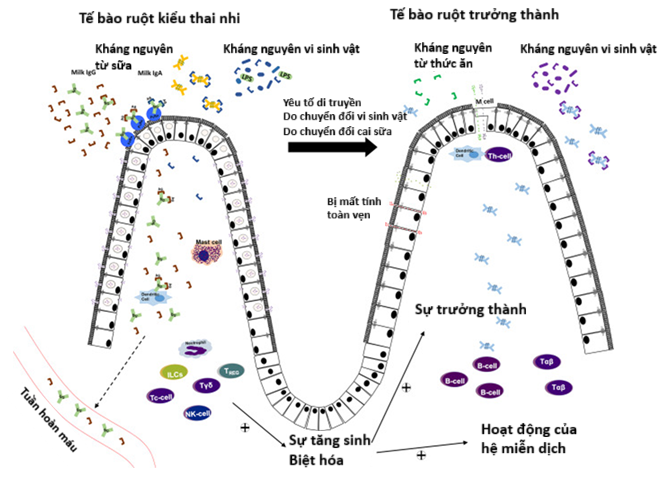 |
| Figure 1. Illustration of the developmental changes in the small intestine, showing the replacement of fetal-type intracellular epithelial cells with mature epithelial cells during prenatal/postnatal development in immature mammals [6]. |
Within the body, high-molecular-weight substances and damaged organelles are engulfed by macrophages, degraded into smaller nutrients, and subsequently supplied to the body. Microorganisms and toxic chemicals are destroyed by reactive oxygen species (ROS) secreted during phagocytosis [7]. Thus, macromolecular food components such as soluble polysaccharides and soluble proteins can still be partially absorbed, contributing to immune stimulation and providing a small nutritional source.
This immunostimulatory mechanism has been extensively studied with β-glucan [2], a compound now widely commercialized as a pharmaceutical product. β-glucans are soluble fibers of large molecular size, generally not absorbed by the digestive system. Nevertheless, oral β-glucan administration has been shown to increase β-glucan concentrations in peripheral blood, with significantly higher levels observed in treated groups compared to controls [5]. This provides a rationale for the immunostimulatory role of β-glucans. Structurally, β-glucans are polymers of D-glucose monomers linked by β-glycosidic bonds. Unlike starch (linked by α-glycosidic bonds), β-glucans can not be digested by the human digestive system. They are present in most plant- and microorganism-derived foods. While natural β-glucans are typically high-molecular-weight or insoluble, in the digestive system, gastric acid and β-glucan-degrading microorganisms can hydrolyze them into shorter-chain structures [8], enabling absorption and immunostimulatory activity in the host.
It has long been recognized that small amounts of soluble proteins absorbed via the gastrointestinal tract can reach peripheral tissues and activate immune responses [5]. The immunostimulatory roles of serum filtrates and dietary proteins are well-documented examples [9]. Dietary proteins also contribute to immune enhancement [4]. This effect is mediated by absorption of small quantities of soluble protein from food, distinct from the traditional nutritional role of proteins in tissue growth and development (which emphasizes amino acid balance and quantity). Foods of both plant and animal origin contain proteins, and therefore all have the potential to stimulate the immune system.
Thus, in food, ingredients that are still considered non-absorbable include starch, protein and soluble fiber that can be partially absorbed by the body and play an important role in stimulating and enhancing the activity of the immune system.
Ngo Duy Sa - Faculty of Food Science and Technology
References
Bounous, G., Létourneau, L., & Kongshavn, P. A. L. (1983). Influence of Dietary Protein Type on the Immune System of Mice. The Journal of Nutrition, 113(7), 1415-1421. https://doi.org/https://doi.org/10.1093/jn/113.7.1415
Chan, G. C., Chan, W. K., & Sze, D. M. (2009). The effects of beta-glucan on human immune and cancer cells. J Hematol Oncol, 2, 25. https://doi.org/10.1186/1756-8722-2-25
clevelandclinic. Inflammation. my.clevelandclinic.org. https://my.clevelandclinic.org/health/symptoms/21660-inflammation
Cockram, T. O. J., Dundee, J. M., Popescu, A. S., & Brown, G. C. (2021). The Phagocytic Code Regulating Phagocytosis of Mammalian Cells. Front Immunol, 12, 629979. https://doi.org/10.3389/fimmu.2021.629979
Hino, S., Nishimura, N., Matsuda, T., & Morita, T. (2020). Intestinal Absorption of β-Glucans and Their Effect on the Immune System. https://doi.org/10.20944/preprints202012.0250.v1
Lee, S. H., Jang, G. Y., Hwang, I. G., Kim, H. Y., Woo, K. S., Kim, K. J., Lee, M. J., Kim, T. J., Lee, J., & Jeong, H. S. (2015). Physicochemical Properties of β-Glucan from Acid Hydrolyzed Barley. Prev Nutr Food Sci, 20(2), 110-118. https://doi.org/10.3746/pnf.2015.20.2.110
Mohamed Abd El-Aziz, T., Garcia Soares, A., & Stockand, J. D. (2019). Snake Venoms in Drug Discovery: Valuable Therapeutic Tools for Life Saving. Toxins (Basel), 11(10). https://doi.org/10.3390/toxins11100564
Weström, B., Arévalo Sureda, E., Pierzynowska, K., Pierzynowski, S. G., & Pérez-Cano, F. J. (2020). The Immature Gut Barrier and Its Importance in Establishing Immunity in Newborn Mammals. Front Immunol, 11, 1153. https://doi.org/10.3389/fimmu.2020.01153
Wu, H., Bao, Y., Wang, X., Zhou, D., & Wu, W. (2017). Alkyl polyglycoside, a highly promising adjuvant in intranasal split influenza vaccines. Hum Vaccin Immunother, 13(6), 1-9. https://doi.org/10.1080/21645515.2016.1278098